Материал для построения клеточной стенки секретирует сам заключенный в ней – протопласт. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку.
Первичная клеточная стенка состоит из целлюлозных мицелл, которые образуют микрофибриллы, погруженных в матрикс, в состав которого входят сложные полисахариды.
Целлюлоза представляет собой полисахарид (полимер глюкозы). В ней заключено около 50 % углерода, находящегося в растениях. 20–40 % материала клеточной стенки составляет целлюлоза. Целлюлозные волокна представляют собой длинные цепи – приблизительно из 10 000 остатков глюкозы. Из каждой цепи выступает наружу множество ОН-групп. Эти группы направлены во все стороны и образуют водородные связи с соседними цепями, что обеспечивает жесткое поперечное сшивание всех цепей. Цепи объединены друг с другом, образуя микрофибриллы, в которых происходит образование пространственных решеток, кристаллоподобных участков, так называемых мицелл (рис. 1).
Рис. 1.
1 – микрофибрилла; 2 – мицелла; 3 – межмицеллярное пространство (диаметр 1 нм); 4 – межмикрофибиллярное пространство (диаметр ~ 10 нм)
Особо важное значение для той роли, которую выполняет целлюлоза в клеточных стенках имеет ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали.
Микрофибриллы, погруженные в матрикс, образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства делят на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях.
Гемицеллюлозы – это смешанная группа полисахаридов, растворимая в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлозы, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Молекулы пектина представляют собой цепочки полигалактуроновой кислоты, свободные карбоксильные группы которой составляют резервуар фиксированных отрицательных зарядов в клеточных стенках.
Отметим также, что размер межмицеллярного и межфибриллярного пространства (пор) составляет соответственно 1 и 10 нм, что значительно превышает размер многих растворенных веществ, транспортируемых в растениях.
Среди вещества матрикса клеточных стенок имеются и белки . Это гликопротеиды, белковая фракция которых сильно обогащена аминокислотой оксипролином. Структурный белок клеточной стенки получил название экстенсина.
Таким образом, свободные карбоксильные группы полисахаридов и белков обеспечивают большое число отрицательных фиксированных зарядов в клеточных стенках. Свободные аминогруппы белков ответственны за меньшее количество положительных зарядов. Заряженные группы матрикса образуют доннановскую систему, играющую важную роль в процессе транспорта ионов в тканях растений. Клеточные стенки гидратированы: 60–70 % их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной стенки отлагаются дополнительные слои целлюлозы, т. е. образуется вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать со вторичным утолщением (вторичным ростом) самого растения, т. е. увеличением толщины ствола в результате добавления новых клеток.
Самый внутренний, очень тонкий последний слой вторичной стенки вследствие особенностей структуры и состава принято называть третичной стенкой. Наряду с целлюлозой и пектином в этом слое имеется много гемоцеллюлоз: в третичной стенке микрофибриллы, по-видимому, образуют сетчатую структуру. Обращенная внутрь клетки поверхность третичной стенки обычно покрыта субмикроскопическими бородавчатыми выростами.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и вторичные) пропитываются лигнином – сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляется контакт между соседними клетками при помощи плазмодесм. Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие. Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. В ряде случаев клетки пропитываются суберином или покрыты восковым налетом – кутикулой.
Клеточная стенка, как упоминалось, пронизана порами, сквозь которые проходят плазмодесмы. Плазмодесма – это не просто нить цитоплазмы, соединяющая протопласты соседних клеток, а органелла имеющая сложное строение. Снаружи плазмодесма покрыта плазмалеммой, которая переходит из одной клетки в другую. Внутри плазмодесмы находится канал (десмотубола), соединяющий эндоплазматическую сеть соседних клеток. Пора, находящаяся в клеточной стенке, в средней части расширена, а на концах сужена, поэтому на концах плазмалемма почти вплотную подходит к десмотуболе. На 1 мм 2 клеточной стенки приходится от 100 тыс. до 50 млн. плазмодесм.
Совокупность протопластов всех клеток, соединенных плазмодесмами, получила название симпласта.
Сравнительно недавно были открыты эктодесмы – очень тонкие цитоплазматические выросты, пронизывающие наружные стенки эпидермальных клеток листа и доходящие до кутикулы. Они подвижны: могут укорачиваться и удлиняться. Эктодесмы, вероятно, выполняют в многоклеточном организме роль рецепторов: воспринимают разного рода сигналы об изменениях в окружающей среде и передают их затем симпласту.
Основные функции клеточной стенки (оболочки) следующие:
1. Оболочки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обуславливают и тургесцентностьклеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает, и в известной мере, регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт) служит путем, по которому передвигается вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи – плазмодесмы. Плазмодесмы объединяют все протопласты в единую систему – симпласт.
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой – кутикулой, состоящей из воскообразного вещества кутина, что снижает потерю воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершению вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок приспособлены для дальнего транспорта веществ по растению.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды и минеральных веществ.
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ. Таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток (клетки-спутники) площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазмалеммы, что повышает эффективность переноса веществ.
10. Благодаря своим катионобменным свойствам клеточная стенка служит резервуаром, накапливающим катионы.
Клеточная стенка (клеточная оболочка) - характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Культивируемые на специальных питательных средах клетки растений, у которых ферментативным путем удаляется стенка, всегда принимают сферическую форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность.
Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту).
Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса.
Скелетным веществом клеточной стенки растений является целлюлоза (клетчатка), представляющая собой бета-1,4-D-глюкан. Это самое распространенное органическое вещество биосферы. Молекулы целлюлозы представляют собой очень длинные неразветвленные цепи, они располагаются параллельно друг другу группами по нескольку десятков и скреплены многочисленными водородными связями. В результате образуются микрофибриллы, которые создают структурный каркас стенки и обусловливают ее прочность. Микрофибриллы целлюлозы видны только в электронный микроскоп, их диаметр равен 10-30 нм, длина достигает нескольких мкм.
Целлюлоза нерастворима и не набухает в воде. Она очень инертна в химическом отношении, не растворяется в органических растворителях, концентрированных щелочах и разведенных кислотах. Микрофибриллы целлюлозы эластичны и очень прочны на разрыв (сходны со сталью). Эти свойства определяют широкое применение целлюлозы и ее продуктов. Мировая продукция хлопкового волокна, состоящего почти целиком из целлюлозы, составляет 1,5*107 тонн в год. Из целлюлозы получают бездымный порох, ацетатный шелк и вискозу, целлофан, бумагу. Качественную реакцию на целлюлозу проводят с реактивом хлор-цинк-йод, целлюлозная клеточная стенка окрашивается в сине-фиолетовый цвет.
У грибов скелетным веществом клеточной стенки является хитин - полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза.
Микрофибриллы погружены в аморфный матрикс, обычно представляющий собой насыщенный водой пластичный гель. Матрикс является сложной смесью полисахаридов, молекулы которых состоят из остатков нескольких различных сахаров и представляют собой более короткие, чем у целлюлозы, и разветвленные цепи. Матричные полисахариды определяют такие свойства клеточной стенки, как сильная набухаемость, высокая проницаемость для воды и растворенных в ней низкомолекулярных соединений, катионообменные свойства. Полисахариды матрикса делят на две группы - пектиновые вещества и гемицеллюлозы.
Пектиновые вещества сильно набухают или растворяются в воде. Они легко разрушаются под действием щелочей и кислот. Простейшими представителями пектиновых веществ являются растворимые в воде пектовые кислоты - продукты полимеризации альфа-D-галактуроновой кислоты (до 100 единиц), связанных 1,4-связями в линейные цепи (альфа-1,4-D-галактуронан). Пектиновые кислоты (пектины) - это более высокомолекулярные (100-200 единиц) полимерные соединения альфа-D-галактуроновой кислоты, в которых карбоксильные группы частично метилированы. Пектаты и пектинаты - кальциевые и магниевые соли пектовых и пектиновых кислот. Пектиновые кислоты, пектаты и пектинаты растворимы в воде в присутствии сахаров и органических кислот с образованием плотных гелей.
В клеточных стенках растений в основном присутствуют протопектины - высокомолекулярные полимеры метоксилированной полигалактуроновой кислоты с арабинанами и галактанами, у двудольных растений в состав цепей галактуронана входит небольшое количество рамнозы. Протопектины нерастворимы в воде.
Гемицеллюлозы представляют собой разветвленные цепи, построенные из остатков нейтральных сахаров, чаще встречаются глюкоза, галактоза, манноза, ксилоза; степень полимеризации 50-300. Гемицеллюлозы химически более устойчивы, чем пектиновые вещества, они труднее гидролизуются и слабее набухают в воде. Гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ (финиковая пальма, хурма). Пектиновые вещества и гемицеллюлозы связаны взаимными переходами. Помимо полисахаридов, в матриксе клеточных стенок присутствует особый структурный белок. Он связан с остатками сахара арабинозы и поэтому является гликопротеидом.
Матричные полисахариды не просто заполняют промежутки между целлюлозными микрофибриллами. Их цепи располагаются упорядоченно и образуют многочисленные связи как друг с другом, так и с микрофибриллами, что значительно повышает прочность клеточной стенки.
Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, или лигнификация происходит в том случае, если в матриксе откладывается лигнин - полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1) флороглюцин и концентрированная хлористоводородная или серная кислота (одревесневшие стенки приобретают вишнево-красную окраску) и 2) сульфат анилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы.
Опробковение, или суберинизация происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров - суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин - аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку является судан III, окраска оранжево-красная.
Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки - кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет.
Минерализация клеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Приобретенная в результате окремнения жесткость стеблей и листьев служит защитным средством против улиток, а также значительно снижает поедаемость и кормовую ценность растений.
У некоторых специализированных клеток наблюдается ослизнение клеточной стенки. При этом вместо целлюлозной вторичной стенки происходит отложение аморфных, сильно гидратированных кислых полисахаридов в виде слизей и камедей, близких по химической природе к пектиновым веществам. Слизи хорошо растворяются в воде с образованием слизистых растворов. Камеди клейкие, вытягиваются в нити. В сухом виде они имеют роговую консистенцию. При отложении слизи протопласт постепенно оттесняется к центру клетки, его объем и объем вакуоли постепенно уменьшаются. В конце концов, полость клетки может целиком заполниться слизью, и клетка отмирает. В некоторых случаях слизь может проходить через первичную клеточную стенку на поверхность. В синтезе и секреции слизи основное участие принимает аппарат Гольджи.
Выделяемая растительными клетками слизь выполняет различные функции. Так, слизь корневого чехлика служит в качестве смазки, облегчающей рост кончика корня в почве. Слизевые железки насекомоядных растений (росянка) выделяют ловчую слизь, к которой приклеиваются насекомые. Слизь, выделяемая наружными клетками семенной кожуры (лен, айва, подорожники), закрепляет семя на поверхности почвы и защищает проросток от высыхания. Слизь окрашивается реактивом метиленовым синим в голубой цвет.
Выделение камедей обычно происходит при поранении растений. Например, камедетечение из пораненных участков стволов и ветвей часто наблюдается у вишни и сливы. Вишневый клей представляет собой застывшую камедь. Камедь выполняет защитную функцию, закрывая рану с поверхности. Образуются камеди в основном у древесных растений из семейств бобовых (акации, трагакантовые астрагалы) и розоцветных подсемейства сливовых (вишня, слива, абрикос). Камеди и слизи используются в медицине.
Клеточная стенка является продуктом жизнедеятельности протопласта. Полисахариды матрикса, гликопротеид стенки, лигнин и слизи образуются в аппарате Гольджи. Синтез целлюлозы, образование и ориентация микрофибрилл осуществляются плазмалеммой. Большая роль в ориентации микрофибрилл принадлежит микротрубочкам, которые располагаются параллельно откладывающимся микрофибриллам вблизи плазмалеммы. Если микротрубочки разрушить, образуются только изодиаметрические клетки.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной, или межклеточной пластинкой (межклеточным веществом) (рис. 16). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными. Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни (рис. 16).
Рис. 16.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной (рис. 17).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 16). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки - ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки - поры. Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов - простые и окаймленные (рис. 17).

Рис. 17. Типы пор: А - клетки с вторичными стенками и многочисленными простыми порами; Б - пара простых пор; В - пара окаймленных пор
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры, или поровой мембраной. В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 18).
Плазмодесмы присущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико - от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр - центральный стержень плазмодесмы, непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.

Рис. 18. Плазмодесмы под электронным микроскопом (схема): 1 - на продольном срезе; 2 - на поперечном срезе; Пл - плазмалемма; ЦС - центральный стержень плазмодесмы; ЭР - элемент эндоплазматического ретикулума
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом, а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
У окаймленных пор (рис. 17) канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время не снижая прочности клеточной стенки.
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.

Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.

Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.
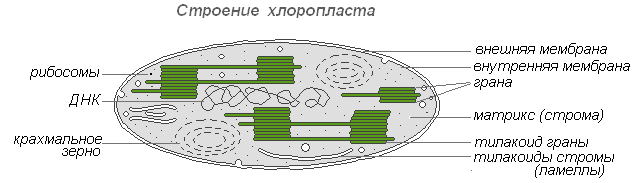
Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.
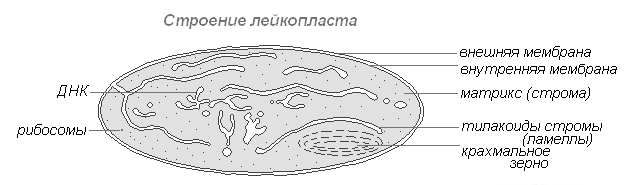
Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.
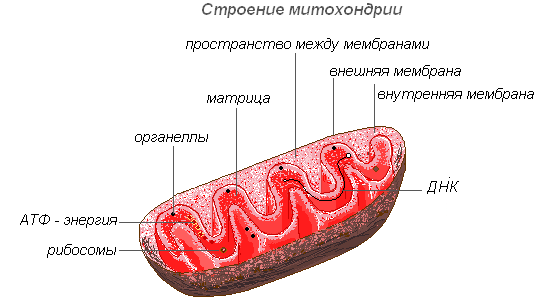
Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.

Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.

Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
КЛЕТОЧНАЯ СТЕНКА, клеточная оболочка одноклеточных, колониальных и многоклеточных грибов и растений. У многоклеточных растений все клеточные стенки и межклетники образуют единую общеорганизменную структуру (апопласт), обеспечивающую передвижение веществ от клетки к клетке. Толщина клеточной стенки варьирует у разных клеток (у растительных, например, от 0,1 мкм до многих десятков мкм). Формирующие её вещества представлены преимущественно полисахаридами. Они образуют матрикс (аморфное вещество) и включённые в него тонкие нити разного размера - микро и макрофибриллы, которые практически нерастяжимы и обеспечивают прочность клеточной стенки на разрыв. Такие микро и макрофибриллы сложены параллельными линейными неразветвлёнными молекулами хитина (у большинства грибов), целлюлозы (у некоторых грибов и большинства растений), ксиланов (у ряда зелёных и красных водорослей), маннанов (у некоторых зелёных водорослей). На их долю приходится от «4% до «90% сухой массы клеточной стенки, что зависит от таксономической принадлежности организма, типа ткани и возраста клетки. Основу матрикса составляют разветвлённые и неразветвлённые молекулы пептидогликанов, маннанов (грибы), пектинов и гемицеллюлоз (высшие растения), альгинатов и фукоидана (бурые водоросли), которые соединены различными связями в ажурную трёхмерную сеть, проницаемую для водных растворов. Матрикс включает также белки (в том числе гликопротеины), роль которых в клеточных стенках плохо изучена.
Образование клеточной стенки начинается с отложения на поверхности плазматической мембраны слоя матрикса, биополимеры которого синтезируются внутри клетки и путём экзоцитоза выделяются на её поверхность. Синтез микро и макрофибрилл происходит из выделяемых протопластом мономерных предшественников с помощью подвижных белковых комплексов. Последние пронизывают насквозь мембрану и связаны с подстилающими её микротрубочками цитоскелета. У одноклеточных и колониальных организмов изначальный слой матрикса лишён микро и макрофибрилл, часто ослизняется; у спор он инкрустируется минеральными веществами и/или полифенолами либо замещается так называемым спорополленином; у многоклеточных организмов изначальный матрикс становится межклеточной пластинкой. В дальнейшем протопласт формирует собственно клеточные стенки, матрикс которой является непосредственным продолжением изначального слоя. У грибов в толще клеточной стенки образуется срединный слой из пептидогликанов без фибрилл. У растений различают первичную и вторичную клеточные стенки. Первая образуется во время роста клетки и завершается вскоре после его окончания; содержит небольшое число беспорядочно ориентированных микро и макрофибрилл, что позволяет стенке растягиваться по мере клеточного роста. Клетки некоторых типов высших растений на первичной клеточной стенке формируют изнутри вторичную клеточную стенку с микро и макрофибриллами, располагающимися плотно, параллельно друг другу, с хорошо выраженной слоистой структурой. Фибриллы различных слоёв вторичной клеточной стенки ориентированы в разных направлениях, вследствие чего она становится нерастяжимой. Клеточные стенки могут подвергаться обызвествлению (например, у «известковых» зелёных и красных водорослей), окремнению (например, у хвощей), одревеснению (у сосудистых растений), опробковению (у высших растений), замещению пектиново-гемицеллюлозного матрикса каллозой (в ситовидных полях элементов флоэмы).
Клеточная стенка выполняет функции внешнего скелета клетки, защитной оболочки, регулятора водного режима, участвует в обменных процессах, передаче сигналов внутрь клетки из окружающей среды. Гемицеллюлозы клеточной стенки в эндосперме семян являются запасным веществом для зародыша (хурма, пальмы). Некоторые клетки высших растений в зрелом состоянии лишаются протопласта и представлены исключительно клеточной стенкой (например, клетки феллемы, проводящие элементы ксилемы); в основном из клеточной стенки отмерших клеток состоит древесина. Фактически именно клеточные стенки служат сырьём для деревообрабатывающей и целлюлозно-бумажной промышленности, производства вискозы, а также получения агара и альгинатов для медицинской, пищевой и косметической промышленности. О клеточной стенке бактерий смотри в статьях Бактерии, Грамотрицательные и грамположительные бактерии.
Наименьшей частью организма является клетка, она способна существовать самостоятельно и имеет все признаки живого организма. В данной статье мы узнаем, какое строение имеет растительная клетка, кратко расскажем об её функциях и особенностях.
Строение клетки растения
В природе существуют как одноклеточные растения, так и многоклеточные. Например, в подводном мире можно встретить одноклеточные водоросли, которые имеют все функции присущие живому организму.
Многоклеточная особь - это не просто набор клеток, а единый организм, способный образовывать различные ткани, органы, которые взаимодействуют друг с другом.
Строение растительной клетки у всех растений одинаковое и состоит из одних и тех же компонентов. Её состав следующий:
- оболочка (пластинка, межклетник, плазмодесмы и плазмолеммы, тонопласт);
- вакуоли;
- цитоплазма (митохондрии; хлоропласты и другие органоиды);
- ядро (ядерная оболочка, ядрышко, хроматин).

Рис. 1. Строение клетки растения.
В отличие от животной, растительная клетка имеет особую целлюлозную оболочку, вакуоль и пластиды.
Изучение строения и функций растительной клетки показало, что:
ТОП-4 статьи которые читают вместе с этой
- самой значительной частью в организме является ядро , которое отвечает за все происходящие процессы. Оно содержит наследственную информацию, которая передаётся из поколения в поколение. От других органоидов отделяет ядро ядерная оболочка;
- бесцветное вязкое вещество, которое наполняет клетку, называется цитоплазмой . Именно в ней находятся все органоиды;
- под клеточной стенкой находится мембрана (тонопласт) , которая отвечает за обмен веществ. Это тоненькая плёнка, отделяющая оболочку от цитоплазмы;
- клеточная стенка достаточно прочная, так как в её состав входит целлюлоза. Поэтому функциями стенки является защита и придача формы;
- маленькими составными компонентами являются пластиды
.
Они могут быть цветными или бесцветными. Так, например, хлоропласты имеют зелёный цвет, именно в них происходит процесс фотосинтеза;
- внутренняя полость, заполненная соком, называется вакуолью . Размер её зависит от возраста организма: чем он старше, тем больше вакуоль. В состав сока входит водный раствор минеральных солей и органических веществ. Он содержит различные сахара, ферменты, минеральные кислоты и соли, белки и пигменты;
- митохондрии способны передвигаться вместе с цитоплазмой, их основная роль - обмен веществ. Именно здесь происходит процесс дыхания и образования АТФ;
- аппарат Гольджи может иметь различные формы (диски, палочки, зёрнышки). Его роль - накопление и выведение ненужных веществ;
- рибосомы синтезируют белок. Находятся они в цитоплазме, ядре, митохондриях, пластидах.
- в отличие от других живых организмов, растения имеют вакуоль, которая хранит все питательные и полезные вещества, расщепляет отжившие старые органеллы и белки;
- клеточная стенка по своему составу отличается от грибного хитина и стенок бактерий. В её состав входит целлюлоза, пектин и лигнин;
- связь между клетками осуществляется при помощи плазмодесм – так называемые поры в клеточной стенке;
- пластиды имеются только в растительном организме. Помимо хлоропластов это могут быть лейкопласты, которые делятся на два вида: одни из них запасают жиры, другие - крахмал. А также хромопласты, которые синтезируют и хранят пигменты;
- в отличие от животного организма, у растительной клетки нет центриолей.

Рис. 2. Изменения размера вакуоли при росте растения.
Клеточное строение растений учёные открыли ещё в XVII веке. Клетки апельсиновой мякоти видны невооружённым глазом, но чаще всего рассмотреть растительный организм можно под микроскопом.

Рис. 3. Строение аппарата Гольджи.
Особенности растительного организма
Исследование разнообразия царства растений выявило такие особенности:
Что мы узнали?
Будучи самой маленькой частью всего организма, клетка может существовать самостоятельно. Она обеспечивает работу различных тканей и жизненно важных органов. Отличительными компонентами от других особей живой природы является строение клеточной стенки, наличие пластид и вакуолей. Каждый органоид имеет свои функции, без выполнения которых невозможно функционирование всего организма в целом.
Тест по теме
Оценка доклада
Средняя оценка: 4.3 . Всего получено оценок: 1130.



